I CAROTENOIDI
Tesi di laurea della Dott.ssa Daniela Gentile
I terpenoidi sono un gruppo di sostanze diffuse soprattutto nel mondo vegetale. Sono idrocarburi altamente insaturi, appartengono alla classe dei terpenoidi.
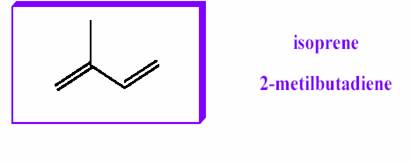
La loro struttura contiene uno scheletro idrocarburico rispondente alla formula bruta (C5H10)n; quindi hanno formula bruta multipla di 5 atomi di C, ossia di una unità C-5, detta unità isoprenica o isoprene.
L’isoprene è un idrocarburo con 5 atomi di carbonio ed una ramificazione; questi si legano tra loro seguendo la legge “testa-coda” e, a seconda di quante unità concorrono a formare i legami, si distinguono in:
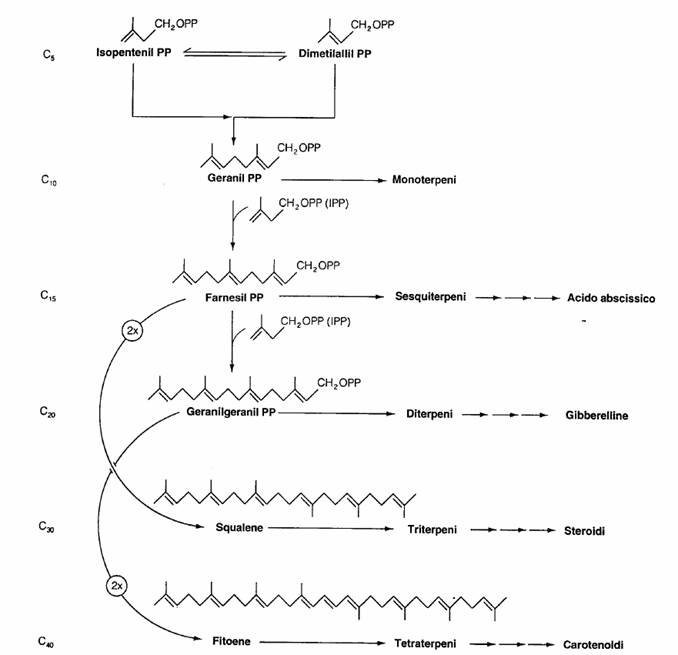
Monoterpeni: sono formati da due unità isopreniche unite testa-coda ed hanno quindi 10 atomi di carbonio.
Sesquiterpeni: sono formati da tre unità isopreniche unite testa-coda ed hanno 15 atomi di carbonio.
Diterpeni: sono formati da quattro unità isopreniche unite testa-coda ed hanno 20 atomi di carbonio.
Triterpeni: sono formati da due unità sesquiterpeniche unite testa-testa ed hanno trenta atomi di carbonio.
Tetraterpeni: sono formati da due unità diterpeniche unite testa-testa ed hanno 40 atomi di carbonio.
Politerpeni: sono formati da n unità isopreniche unite testa-coda.
La classificazione si basa sul contenuto di atomi di carbonio e sulla presenza o meno di strutture cicliche.
Strettamente correlati con i tetraterpeni sono i carotenoidi, un gruppo di pigmenti, di colore dal giallo al violetto, molto diffusi in natura, privi di azoto nella molecola e solubili nei grassi.
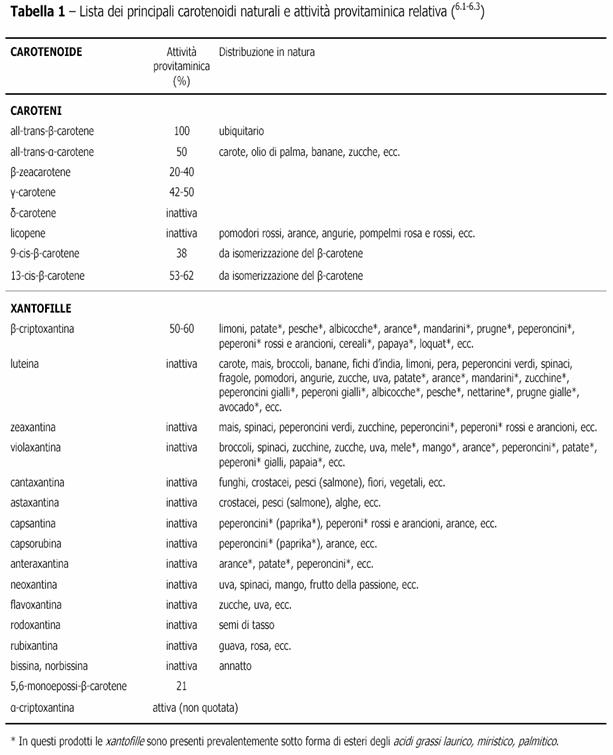
Lo scheletro della loro molecola consiste generalmente di una porzione centrale, con 22 atomi di carbonio, e due terminali di 9 atomi di carbonio ciascuna. Le unità terminali possono essere acicliche, come nel licopene, oppure tutte e due cicliche, come nell’ α e β-carotene, o una ciclica e l’altra aciclica, come nel γ-carotene . Possono inoltre presentare una ampia varietà di gruppi, per esempio alcolici, chetonici, epossidici, benzenici ecc..
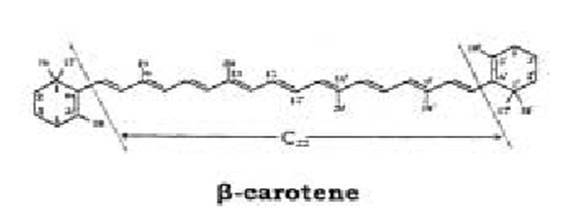
Per maggiore chiarezza, la struttura molecolare dei carotenoidi viene rappresentata trascurando gli atomi di idrogeno e non riportando il simbolo del carbonio, ma soltanto i legami che uniscono tra loro i vari atomi.
Alla famiglia dei carotenoidi appartengono:
1) CAROTENI: carotenoidi non ossigenati a struttura idrocarburica dienica come ad es. β-carotene, α-carotene, licopene e altri; a loro volta i caroteni possono essere classificati in aciclici (es. licopene), monociclici (es. γ-carotene) e biciclici (es. α e β-caroene);
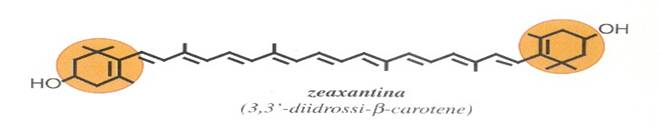
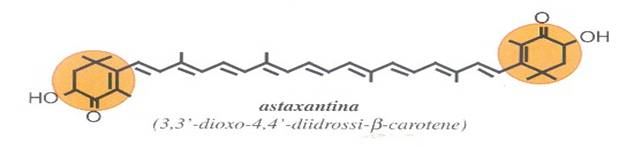
2) XANTOFILLE: derivati ossigenati dei caroteni come ad es. luteina, zeaxantina, β-criptoxantina, astaxantina, bissina, capsantina, e altre; nei vegetali le xantofille possono trovarsi in forma libera o esterificate con acidi grassi.
La posizione e il tipo dell'aggruppamento ossigenato non sono sempre gli stessi.
Nella zeaxantina l'ossigeno è incorporato in un aggruppamento formato dall'unione di un ossigeno con un idrogeno (ossidrile, -OH), mentre nella astaxantina, a fianco di due analoghi ossidrili, si trova un atomo di ossigeno legato al carbonio con un legame doppio (carbonile, C=O). Le posizioni più frequentemente occupate dall'ossigeno sono identificate, nella nomenclatura ufficiale, con i numeri 3, 3', 4, 4' .
La catena comune ai carotenoidi contiene una serie di legami doppi alternati, questa alternanza è chiamata coniugazione e conferisce al sistema particolari proprietà ottiche. Quindi, il "cromoforo" dei carotenoidi è costituito dalla lunga catena formata da legami doppi alternati a legami semplici, ed è in grado di assorbire alcune radiazioni della luce visibile (cioè alcuni colori) riflettendo i colori complementari a quelli assorbiti. Aumentando la lunghezza della coniugazione, il colore riflesso si sposta dal giallo al violetto, la spiegazione sta nel numero di doppi legami tra gli atomi di carbonio C=C .
Tali sostanze hanno una banda d’assorbimento della luce visibile molto ampia che presenta 3 picchi in corrispondenza del verde-violetto. Il picco massimo d’assorbimento dipende dalla costituzione chimica: ogni doppio legame tra gli atomi di carbonio in più sposta il picco massimo verso il rosso di circa 0,02 mm quando il pigmento è disciolto in carbonio disolfuro.
Il legame doppio contenuto nel gruppo terminale non è necessariamente in posizione alternata rispetto ai legami doppi della catena, ma può essere anche più lontano, cioè può non essere coniugato con la catena. Il gruppo terminale con il legame doppio coniugato è chiamato β; mentre il gruppo terminale con il legame doppio non coniugato è chiamato ε. ε,ε-carotenoidi (xantofille) appaiono di un colore giallo limone, β,ε-carotenoidi (luteina) giallo oro, β, β-carotenoidi (β-carotene, β-criptoxantina, zeaxantina) arancione.
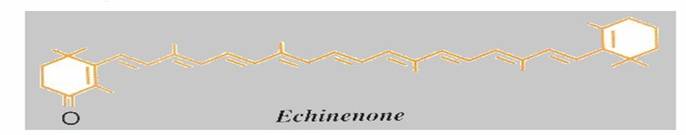
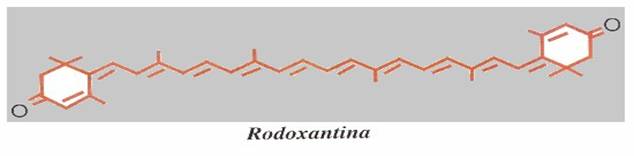
La presenza di un carbonile (C=O chiamato anche gruppo OXO) in posizione adiacente all'ultimo legane doppio (posizione 4) aumenta la coniugazione di una unità, per cui un 4-oxo-β,β -carotenoide (echinenone) sarà arancione rosso e un 4, 4'-dioxo-β,β-carotenoide (cantaxantina, astaxantina), decisamente rosso. La rodoxantina, la cui molecola contiene ben 14 legami doppi coniugati appare di un bel colore rosso-violetto.
Secondo la nomenclatura sistemica le molecole dei carotenoidi vengono considerate divise in due parti e ogni composto si fa derivante dal carotene. I due gruppi terminali sono descritti con lettere greche mentre prefissi e suffissi convenzionali descrivono i livelli di idrogenazione e la presenza di gruppi sostituenti.
I doppi legami non ciclici sono sempre nella configurazione trans, anche se per esposizione alla luce e ad altri agenti, i carotenoidi naturali possono trasformarsi in una miscela di stereoisomeri (isomerizzazione trans-cis), nei quali uno o più doppi legami si sono spostati in posizione cis.
Per il β-carotene, ad esempio, è stata ipotizzata l’esistenza di 272 isomeri, anche se poi in realtà il numero di quelli che si ottengono, per ragioni di ingombro sterico, è minore. Per il licopene sono state ritrovate in natura, in alcune varietà di pomodoro, nelle alghe e nei fiori di calendula, una ventina di queste forme poli-cis., che sono state denominate prolicopeni.
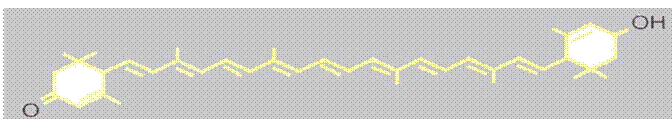
Astaxantina


I carotenoidi cristallizzano facilmente; inoltre, quando sono in soluzione, e specialmente in presenza di luce, vanno incontro a ossidazione e si trasformano in composti incolori. Sono invece molto stabili quando la pressione d’ossigeno è bassa, al buio e a basse temperature.
Sono insolubili in acqua, solubili nei grassi, in etere e acetone.
Un tempo i carotenoidi degli animali erano detti lipocromi, poiché erano stati scoperti dissolti nei grassi.
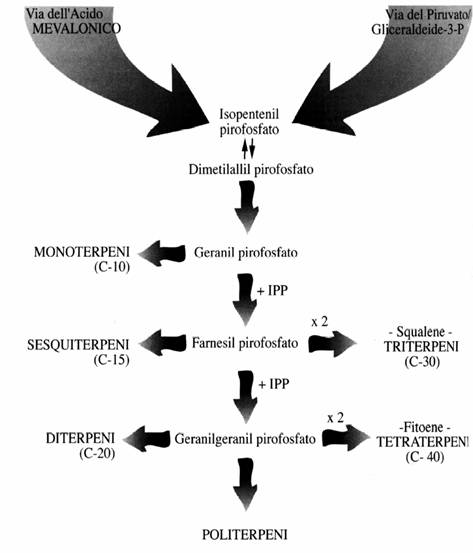
Sintesi dei Carotenoidi
1) Via dell’acido mevalonico a formare l’isopentenil pirofosfato (IPP).
2) Sintesi del geranilgeranil pirofosfato (GGPP).
3) Sintesi dei vari carotenoidi (β-carotene etc.)
Sintesi dell’acido mevalonico
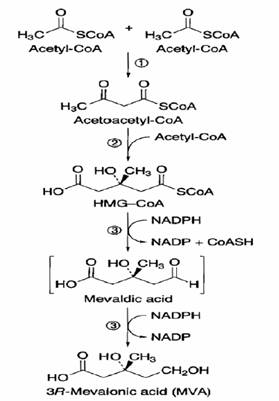
2 Acetil-CoA ---------------> Acetoacetil-CoA
Reazione catalizzata dalla acetil-CoA acetiltransferasi
Acetocetil-CoA + Acetil-CoA ----------> Idrossimetilglutaril-CoA (HMG-CoA)
Reazione catalizzata dalla HMG-CoA sintasi
HMG-CoA + NADPH----------- > Ac. Mevaldico + NADP + CoA-SH
Ac. Mevaldico + NADPH -------> Ac. Mevalonico NADP
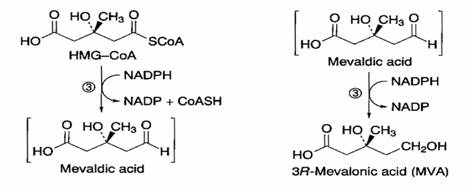
La reazione (divisa in due steps) e’ catalizzata dalla HMG-CoA reduttasi. Enzima localizzato a livello della membrana plasmatica.
La seconda reazione (Ac mevaldico ------------> Ac. mevalonico) regola la velocita’ globale della via dell’acido mevalonico.
Sintesi dell’ unità isoprenica attiva a C5
(Isopentenil-PP).
Acido mevalonico + ATP à Mevalonato 5-P
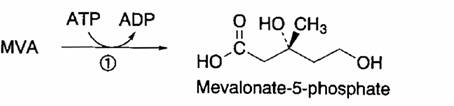
Reazione catalizzata dalla mevalonato 5- P chinasi.
L’enzima risulta localizzato prevalentemente nel citosol.
Mevalonato 5-P + ATP à Mevalonato 5
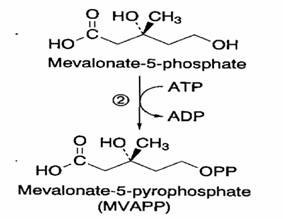
Reazione e’catalizzata dalla mevalonato 5- P chinasi. L’enzima risulta localizzato prevalentemente nel citosol.
Mevalonato 5-PP + ATP à IPP
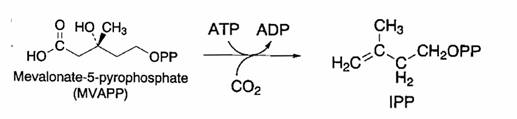
Reazione e’catalizzata dalla mevalonato 5 difosfato decarbossilasi. L’enzima richiede ATP e cationi bivalenti come cofattori.
Sintesi delle principali classi dei terpenoidi
Le reazione comune che e’ alla base della sintesi di tutte le classi dei terpenoidi e’ la seguente:
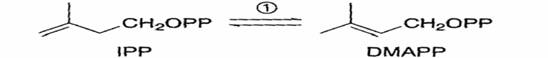
La reazione e’ catalizzata dall’isopentenil pirofosfato isomerasi
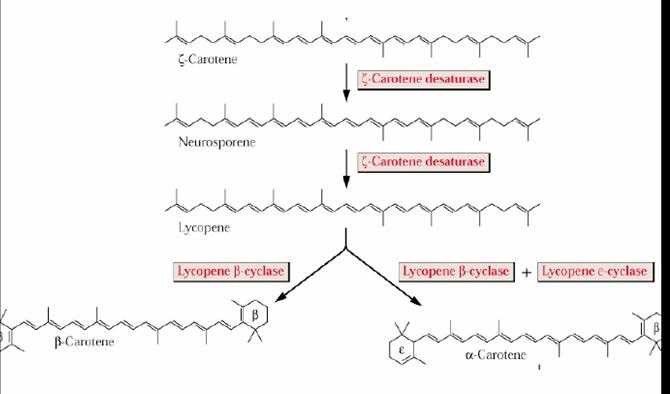
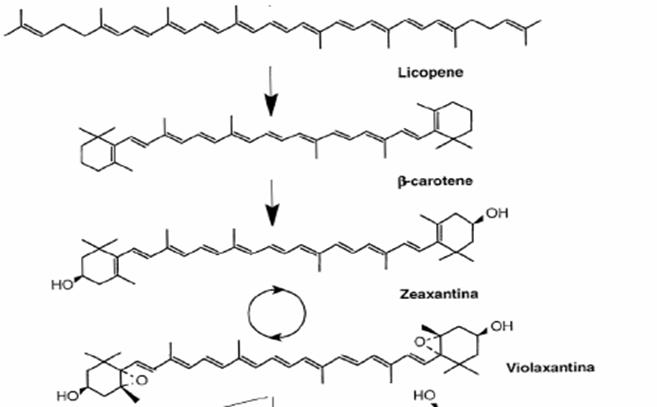
Il primo passaggio della biosintesi dei carotenoidi implicano la condensazione di due molecole di GGPP ad opera della fitogene sintasi (PSY) che si compie in due tappe portando prima alla formazione di prefitoene pirofosfato(PPPP) e quindi di fitoene. La reazione di condensazione porta alla perdita dell’idrogeno e del gruppo pirofosforico in C-1’ della stessa molecola. Quindi si ha il distacco del gruppo pirofosforico in c-1 del PPPP seguito da un riaggiustamento 1-1’ con la conseguente formazione del fitogene, che può risultare 15-cis oppure tutto in trans. Successivamente una serie di quattro desaturazioni (ad opera di fitoene desaturasi) portano alla formazione dei doppi legami coniugati convertendo il fitoene (incolore) in fitofluene (incolore) in ς-carotene (giallo), in neurosporene (arancio) e licopene (rosso).
L’azione della licopene ciclasi porta alla formazione dei carotenoide biciclico β-carotene.
Le xantofille sono degli idrossi, epossi, ossi, furanossi derivati dei caroteni che si formano negli ultimi stadi della via biosintetica.
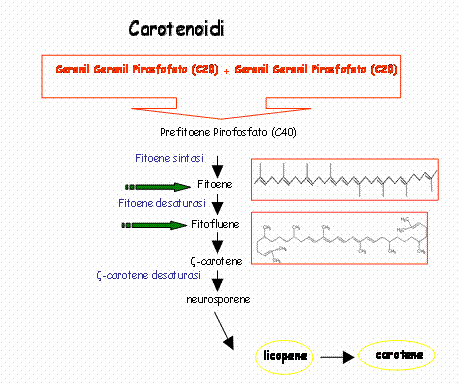
Le prime xantofille, come la zeaxantina, si formano per ossidrilazione degli anelli iononici in posizione 3 e 3’, l’ulteriore epossidazione nelle posizioni 5,6 e 5’,6’ porta alla formazione di xantofille come la violaxantina. Quest’ultima reazione, l’epossidazione, può avvenire in presenza di luce portando all’interconversione della violaxantina nella zeaxantina e viceversa.
Distribuzione in natura
Attualmente, esistono oltre 600 tipi di carotenoidi conosciuti, e nonostante la loro simile composizione chimica, posseggono nomi assai diversi, correlati alla fonte biologica da cui sono stati isolati per la prima volta, dalla quale richiamano la denominazione botanica (carotene da Daucus carota, nelle carote, licopina da Lycopersicum esculentum, nei pomodori, zeaxantina da Zea mays, nel mais…..).
I carotenoidi sono stati isolati dalle piante, o da altri organismi fotosintetici, come le alghe ed alcune specie di funghi e di batteri.
Sono stati ritrovati anche in molti organismi animali, che non sono però in grado di sintetizzare questi composti, e devono assumerli dal mondo vegetale, attraverso l’alimentazione.
Sono responsabili dei magnifici colori di molti frutti e fiori, come pure dei colori di molti uccelli, insetti ed animali marini per i quali possono avere anche uno scopo ornamentale legato al corteggiamento.
Colorano perfino il tuorlo delle uova e il burro, infatti tale colorazione dipende molto dal tipo di alimentazione dell’animale produttore; più i mangimi sono ricchi di carotenoidi, più tali colori saranno intensi.
Nell’organismo animale sono talvolta presenti sotto forma di composti proteici o lipidici che ne mascherano il colore (mimetismo animale); il riscaldamento distruggendo la porzione temolabile della molecola, può invece evidenziarne nuovamente la colorazione (gamberi cotti che diventano rossi).
La luce favorisce la deposizione dei carotenoidi nei tessuti animali: il gamberetto d’acqua dolce, tenuto al buio, si scolorisce; granchi, aragoste e attinie si colorano in proporzione alla luce che ricevono. Se tenuti in soluzione, i carotenoidi vengono scoloriti dall’ossigeno e dalla luce.
Spesso le piante contenenti carotenoidi, come la Bixa orellana e il Crocus sativus (zafferano) ad esempio, sono utilizzate in campo alimentare per la peculiare brillantezza dei colori che da esse si possono ottenere.
La solubilità nei grassi fa della bixina (il carotenoide estratto dai semi della Bixa orellana), classificata dalla legislazione italiana vigente con la sigla E 160 b, un colorante molto usato per ravvivare il colore naturale di oli, burro, margarina e formaggi.
I carotenoidi sono usati largamente come coloranti naturali non tossici e trovano il loro massimo impiego nei cibi, bevande, dolci cosmetici ed alimenti per animali.
Alcuni prodotti della degradazione dei carotenoidi sono importanti fragranze chimiche e vengono utilizzate nell’industria dei profumi. Molti di loro, infatti sono responsabili della profumazione dei fiori e frutti.
Nella pianta sono presenti oltre che nei frutti e nei fiori anche nei cloroplasti dei tessuti verdi (foglie, steli e erba) dove il loro colore viene mascherato da quello della clorofilla, che affiancano nel processo fotosintetico, generalmente complessati con proteine. Sono piuttosto rari nelle radici, anche se ci sono notevoli eccezioni, come nel caso delle carote dove sono presenti in grandi quantità. Anche alcuni semi sono colorati dai carotenoidi, come nel caso del mais che deve il suo colore alla zeaxantina.
Hanno un rilevante ruolo nella protezione delle piante dalla grande produzione di radicali liberi che si verifica durante il processo fotosintetico.
In generale, gli stessi carotenoidi sono stati isolati dalle foglie di tutte le specie vegetali e si è avuta la seguente composizione percentuale: β-carotene (25%-30% del totale), luteina ( circa il 40%), violaxantina (circa il 15%) e neoxantina ( circa il 15%); sono inoltre presenti, anche e in piccole quantità, α-carotene, α- e β-criptoxantina, zeaxantina, anteraxantina e luteina-5,6-epossido. La composizione quantitativa dei carotenoidi nella maggior parte delle foglie è simile, ma può cambiare se le piante sono in condizioni di stress.
Il β-carotene è il tipo predominante nella maggior parte delle foglie verdi, e tanto più elevata è la sua concentrazione, tanto più intensa appare la colorazione delle parti che lo contengono.
Fotosintesi
Processo mediante il quale, a partire da anidride carbonica e acqua e con l’utilizzazione della luce solare quale fonte di energia, vengono prodotti glucosio e ossigeno. Quindi l’energia solare viene trasformata in energia chimica, che è rappresentata dall’energia dei legami chimici delle molecole ottenute con questo processo.
Il prodotto organico della fotosintesi è il glucosio (C6H12O6). Il carbonio e l'ossigeno da convertire in sostanza organica sono forniti dall'anidride carbonica (CO2) atmosferica, e l'idrogeno dall'acqua (H2O). Si tratta di invertire il senso della combustione del glucosio e occorreranno 686 Kilocalorie/mole di glucosio prodotto.
L'equazione chimica che riassume il processo è:
6 CO2 + 6 H2O + 686 Kilocalorie/mole → C6H12O6 + 6 O2
Nella fotosintesi si distingue una “fase luminosa” e una “fase oscura”; la prima comprende l’insieme delle reazioni che richiedono luce, la seconda quelle che non ne necessitano.
Nel corso della prima fase, clorofille e carotenoidi, situati nelle membrane dei cloroplasti, assorbono luce solare: gli elettroni stimolati raggiungono un livello energetico superiore e trasferiscono energia a una molecola di pigmento speciale detta centro di reazione.
La fase oscura si svolge nello stroma e porta alla riduzione dell’anidride carbonica con formazione di composti organici. In tale processo (ciclo di Calvin), uno zucchero a 5 atomi di carbonio, il ribulosio-1,5-difosfato (RuDP), si combina con l’anidride carbonica per formare due molecole di un composto a tre atomi di carbonio, il 3-fosfoglicerato (PGA).
In un giro del ciclo si ha la riduzione di una molecola di CO2; in tre giri del ciclo si forma una molecola di gliceraldeide- 3-fosfato. Due molecole di gliceraldeide-3-fosfato possono combinarsi per formare una molecola di glucosio.
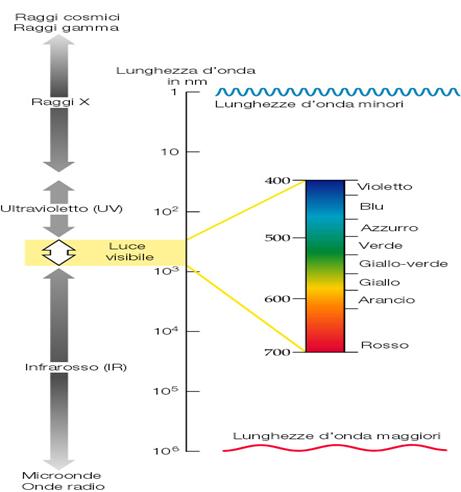
La luce è una piccola porzione dello spettro elettromagnetico radiante che comprende i raggi X, i raggi ultravioletti, le onde radio, le microonde.
Dal punto di visto fisico la radiazione è costituita da particelle dette quanti o fotoni che si diffondono sia sotto forma di onde che di corpuscoli.
I vari tipi di radiazioni differiscono l’una dall’altra per l’energia che contengono e per la lunghezza d’onda (distanza tra un picco e l’altro) che si misura in nm (nanometri) a partire da quelle a cortissima lunghezza d’onda (raggi gamma e raggi X) fino a quelle ad elevata lunghezza d’onda (onde radio).
Il nostro occhio è in grado di percepire solo alcune radiazioni con determinate lunghezze d’onda che vanno da circa 390 nm (blu) fino a circa 760 nm (rosso): questo intervallo viene definito spettro del visibile o luce visibile.
A seconda della sua lunghezza d’onda si hanno diversi tipi di colori della luce.
Sostanze diverse assorbono lunghezze d’onda diverse dello spettro elettromagnetico.
I PIGMENTI sono sostanze che assorbono specifiche lunghezze d’onda ed hanno un colore specifico.

I pigmenti fotosintetici
L’energia luminosa viene assorbita mediante i pigmenti fotosintetici che si trovano nelle piante all’interno di organuli detti cloroplasti.
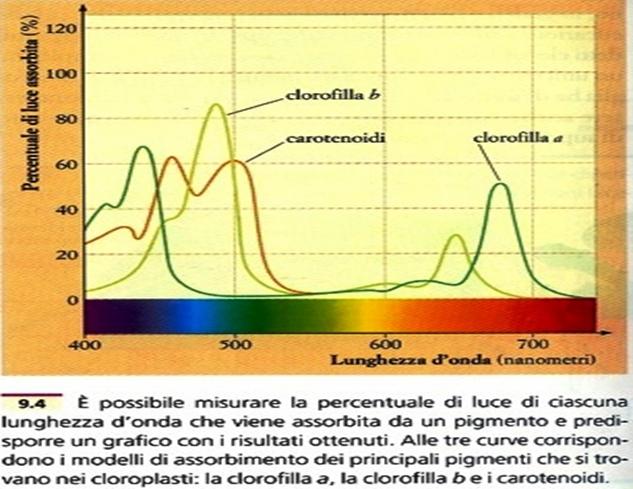
Il pigmento più importante è la clorofilla. Esistono diversi tipi di clorofilla contrassegnati dalle lettere a, b, c, d, variamente distribuite nei vegetali.
L’unica responsabile della conversione di energia luminosa in energia chimica è la clorofilla a, ed è chiamata centro di reazione, mentre gli altri pigmenti che hanno soltanto la funzione di captazione dell’energia sono dette molecole antenna, o pigmenti coadiuvanti.
La clorofilla è costituita da un anello porfirinico con al centro un atomo di Mg2+; a questo anello è attaccato una lunga coda formata da venti atomi di carbonio che è un alcool detto fitolo altamente idrofobo (permette l’inserimento nella membrana lipidica).
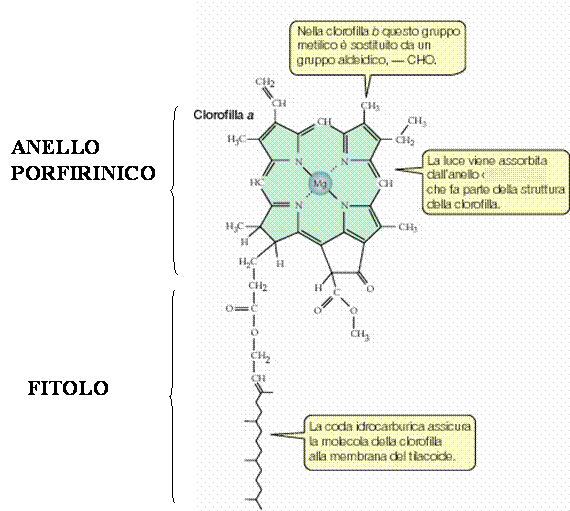

La clorofilla b differisce dalla a soltanto per il gruppo in posizione 3, che nella prima è il gruppo aldeidico(CHO), mentre nella seconda è il gruppo metilico(CH3).
La clorofilla b è di colore verde-grigiastro, e la clorofilla a verde-bluastro. Gli spettri di assorbimento della luce nelle 2 clorofille differiscono, in quanto nella clorofilla a l’assorbimento nella zona del rosso è compresa fra 680 e 640 nm, con un massimo di a 662 nm; mentre nella clorofilla b è spostato fra655 e 630 nm, con un massimo a 642 nm. L’assorbimento nella zona dell’azzurro inizia a 480 nm nella clorofilla b, e per la clorofilla a, a 445 nm.
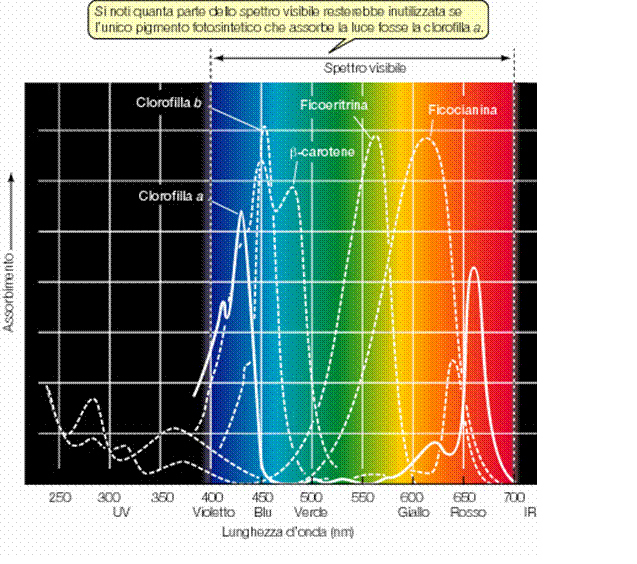
I carotenoidi fanno parte del complesso antenna e sono i pigmenti accessori presenti in tutti gli organismi foto sintetici, mentre le ficobiliproteine pigmenti accessori presenti solo nelle alghe rosse.
I carotenoidi sono composti notevolmente più stabili delle clorofilla, le quali hanno un tempo di degradazione assai rapido. Hanno la funzione di estendere l’assorbimento della luce a regioni dello spettro non coperte in modo adeguato dalle clorofille (da 440 a 470 nm, blu-violetto), per trasmettere poi l’energia alla clorofilla a.
Proteggono inoltre le clorofille da possibili fotossidazioni distruttive che potrebbero verificarsi ad alte intensità luminose.
In assenza del normale corredo di carotenoidi, la pianta muore per fotoossidazione e, ad esempio, molti erbicidi agiscono proprio inibendo la biosintesi dei carotenoidi.
In autunno, quando le piante si preparano a perdere le foglie, il contenuto di clorofilla diminuisce, i carotenoidi manifestano la loro presenza, e le foglie assumono i colori caldi e vivaci caratteristici della stagione.
In alcuna piante, specie nelle conifere, in alcune stagioni o stadi di sviluppo della pianta, nelle foglie o negli aghi si accumulano carotenoidi non comuni come cheto-carotenoidi o seco-carotenoidi. Questi composti non si trovano complessati con le proteine, bensì in goccioline di olio.
I carotenoidi si trovano anche nei tessuti non fotosintetici delle piante (fiori e frutti), situati all’interno dei cromoplasti.
Non sono presenti nei frutti acerbi, ma si formano durante il processo di maturazione, in presenza di ossigeno, esibendo le caratteristiche colorazioni giallo, arancio e rosso.
Nei fiori sono stati osservati 3 pattern principali di carotenoidi:
Ø Quelli che contengono principalmente carotenoidi comuni
Ø Quelli che contengono 5,6- epossicarotenoidi e 5,8-epossicarotenoidi
Ø Quelli che contengono carotenoidi insoliti come l’escoltzaxantina in Escoltzia californica.
I carotenoidi possono essere presenti in tutte le parti anatomiche del fiore, come petali, sepali, stami e foglie.
Nei frutti sono stati identificati più pattern di carotenoidi.
Ø Quelli che contengono principalmente carotenoidi comuni
Ø Quelli che contengono principalmente grandi quantità di licopene e i suoi idrossi derivati (pomodoro)
Ø Quelli che contengono principalmente grandi quantità di β-carotene ed i suoi idrossi derivati (pesche)
Ø Quelli che contengono principalmente miscele di 5-,6- o 5-,8-epossicarotenoidi
Ø Quelli che contengono principalmente grandi quantità di apo-carotenoidi (carotenoidi degradati)
Ø Quelli che contengono principalmente carotenoidi specifici o insoliti (capsantina nel Capsicum annum)
Ruolo dei carotenoidi nell’organismo umano
Gli animali, così come l’uomo non sono in grado di sintetizzare i carotenoidi de novo come fanno i vegetali e devono procurarsi questi micronutrienti esclusivamente attraverso l'alimentazione.
Dei più di 600 carotenoidi naturali conosciuti, solo una ventina si ritrovano nei tessuti e nel plasma umano; tra questi ci sono il licopene, il β-carotene, la luteina e la zeaxantina.
Questi composti sono necessari per un miglior funzionamento dell’ organismo.
Hanno diverse funzioni biologiche molto complesse che si svolgono in più direzioni.
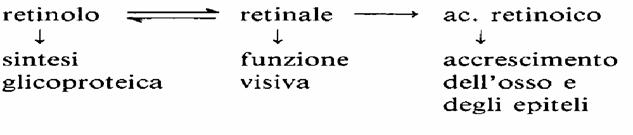
· vengono metabolizzati per produrre vitamina A e acido retinico
· Riducono la genotossicità di molti inquinanti endogeni ed esogeni
· Riducono la presenza delle lesioni precancerose nell’uomo
· Modificano la risposta immunitaria nei mammiferi.
· proteggono le cellule dell’organismo dai dannosi attacchi dell’ambiente esterno (per es. "radicali liberi")
· svolgono funzioni disintossicanti
· sono coinvolti nel processo della vista
· proteggono la pelle dai danni provocati dai raggi ultravioletti
· aumentano la fertilità negli animali
Oltre a queste attività è stato anche dimostrato come i carotenoidi siano in grado di influenzare la comunicazione intercellulare, di rivestire un ruolo importante nei processi infiammatori inibendo le lipoossigenasi ed infine di potenziare il sistema auto-immune, soprattutto nelle manifestazioni patologiche che lo vedono talvolta notevolmente ridotto nella sua funzionalità.
Una loro carenza comporta un aumento del rischio di patologie infiammatorie croniche tra cui l’artrite reumatoide.
Studi
epidemiologici hanno evidenziato una forte correlazione fra la carenza di
carotenoidi e vari tumori dei tessuti epiteliali: polmone, collo dell’utero,
apparato gastrointestinale ed infine la pelle che non è sufficientemente
protetta dai radicali liberi e quindi anche dai raggi solari.
Tanto più elevato è il consumo di carotenoidi, tanto più basso sarà il rischio
di cancro, l’invecchiamento e il cancro hanno delle caratteristiche in comune,
fra cui il rapporto col danno da radicali liberi.
E’ stato, infatti, dimostrato come questi cromofori siano in grado di neutralizzare e di impedire la eccessiva formazione di radicali liberi prodotti dal nostro organismo in quantità sempre maggiore a causa della massiccia presenza di inquinanti ambientali, oltre che per la sovraesposizione ai raggi UV.
Carotenoidi come pro-vitamina A
Degli oltre 600 carotenoidi individuati solo 50 di essi sono precursori di vitamina A, detti “carotenoidi pro-vitamina A”.
Per provitamine A si intendono tutti i carotenoidi che possono essere metabolizzati dall’organismo a retinoidi e che pertanto rappresentano dei precursori della Vitamina A: tra questi, la provitamina A più attiva e conosciuta è il β-carotene (forma all-trans), anche se esistono molti altri carotenoidi che possiedono attività provitaminica A.
 Questi
sono largamente distribuiti nel regno vegetale, il numero di carotenoidi
naturalmente presenti nei prodotti vegetali (frutta, verdura, cereali, ecc.) è
molto elevato e non tutti i carotenoidi possiedono attività provitaminica A;
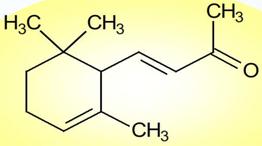
solo quelli che abbiano come gruppo terminale almeno un anello insaturo di
α o β-ionone.
Questi
sono largamente distribuiti nel regno vegetale, il numero di carotenoidi
naturalmente presenti nei prodotti vegetali (frutta, verdura, cereali, ecc.) è
molto elevato e non tutti i carotenoidi possiedono attività provitaminica A;
solo quelli che abbiano come gruppo terminale almeno un anello insaturo di
α o β-ionone.
α-ionone
β-ionone
Tali precursori della vitamina A possiedono attività provitaminica diversa tra loro: le provitamine A diverse dal β-carotene, come ad es. α-caroene (ad es. in carote e zucche), cis-β- caroeni (da isomerizzazione termica/fotochimica del β-carotene), β-criptoxantina (ad es.in mandarini, pesche, papaie) presentano un’attività provitaminica inferiore di circa il 50% rispetto a quella assegnata al β-carotene.
I frutti e i vegetali di colore arancio, come carote, albicocche, meloni e mango, sono di solito i più ricchi di carotenoidi provitamina A, e la loro quantità è direttamente correlata all’intensità della colorazione.
Quelli giallo-arancio contengono sempre β-carotene, ma soprattutto altri carotenoidi provitamina A, mentre in quelli gialli sono presenti innanzitutto xantofille, ad attività provitaminica minore.
I frutti e i vegetali di colore rosso porpora, come pomodori, cavolo rosso e frutti di bosco, contengono carotenoidi e anche grandi quantità di altri pigmenti, per esempio i flavonoidi, che non hanno attività provitaminica A.
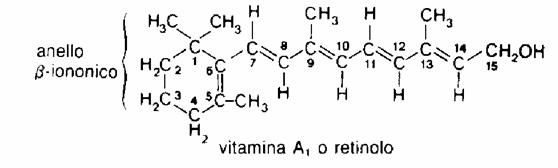
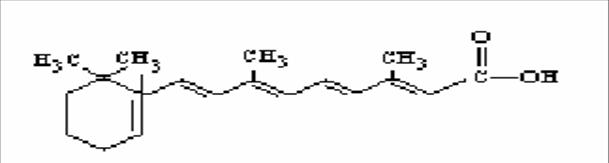
La vitamina A è una delle quattro vitamine liposolubili , in natura si trova in quattro forme diverse: retinolo (vitamina A1), 3-deidroretinolo (vitamina A2), retinale, acido retinoico. Le prime tre forme inoltre (vitamina A1, vitamina A2 ed il retinale) sono interconvertibili tra loro mentre la conversione da retinale ad acido retinoico è irreversibile.
Il retinolo chiamato anche vitamina A1 è un alcol primario caratterizzato da un anello cicloesenico detto β-ionone e da una catena laterale a undici atomi di carbonio ricca di doppi legami che danno origine a vari isomeri geometrici facilmente interconvertibili.
Il 3-deidroretinolo o vitamina A2 è la forma deidrogenata del retinolo, presenta quindi un anello maggiormente insaturo.
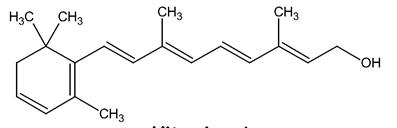
vitamina A2 o 3-deidroretinolo
La A2 ha il 60% di attività biologica in meno rispetto alla vitamina A1, alla quale si trova generalmente associato nei tessuti di pesci di acqua dolce.
Il retinale e l’acido retinoico sono i prodotti di ossidazione del retinolo sulla funzione alcolica prima ad aldeide e poi ad acido carbossilico.
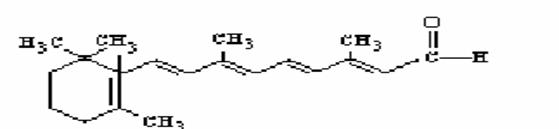
retinale
acido retinoico
La presenza di 4 doppi legami nella catena laterale e’ compatibile con isomerizzazioni cis-trans; esistono quindi diversi stereoisomeri della vitamina A, sebbene solo alcuni siano stericamente possibili; le forme trans sono le più frequenti.
Il retinolo e i suoi prodotti sono composti idrofobici, altamente instabili in presenza di ossigeno, in ambiente acido e alla luce e possono produrre una miscela di svariati prodotti di degradazione.
Le vitamine A1 e A2 sono presenti in natura nei tessuti animali, (fegato, albume d’uovo e latte e derivati) sotto forma di alcooli liberi o esterificati con acido acetico o palmitico , ma anche stearico ed oleico, mentre nei vegetali troviamo i carotenoidi.
I carotenoidi introdotti con la dieta vengono assorbiti dall’intestino dove vengono convertiti in vitanima A, e questa, insieme a quella assunta come tale, per la maggior parte (oltre il 90% di quella contenuta in tutto l’organismo) viene esterificata con acidi grassi a catena lunga, dopo di che viene immessa nella linfa e trasportata al fegato, dove viene immagazzinata; la quota restante non viene esterificata. Il fegato è un deposito di vitamina A disponibile per un lungo periodo; se l’organismo necessita di quantità maggiori di vitamina, gli esteri sono scissi, e il retinolo secreto nel sangue, dove è trasportato complessato con una proteina.
I caroteni provitamina A, sono trasformati in vitamina A a livello della mucosa intestinale per mezzo di particolari reazioni enzimatiche
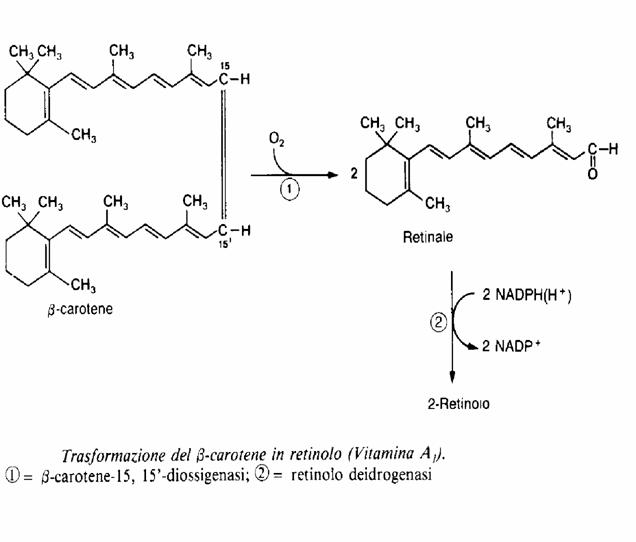
I carotenoidi hanno la struttura di un dimero costituito da 2 unità di retinoide.
Teoricamente da una molecola di β-carotene si ottengono due molecole di retinolo per azione successiva della β-carotene-15, 15’diossigenasi e del retinolo deidrogenasi.
L’intoduzione di una molecola di O2 in corrispondenza del doppi legame 15, 15’ forma inizialmente un perossido in corrispondenza del quale si ha la rottura della molecola, e possedendo due anelli β-iononici, una molecola di β-carotene forma 2 molecole di retinale, che viene successivamente ridotto ad alcool (vitamine A1 e A2). In realtà un equivalente di β-carotene esplica un azione inferiore a quella dei due equivalenti di retinale, sia perché il suo assorbimento intestinale è incompleto, sia perché sia cellule intestinali che gli epatociti, le uniche che contengono β-carotene-15, 15’diossigenasi, non lo convertono quantitativamente in retinolo. Fra l’altro la parte del retinale formatosi viene ossidato nell’intestino ad acido retinoico.
La conversione in vitamina A di un carotenoide precursore dipende da diversi fattori: il livello di vitamina A nell’organismo, le condizioni delle proteine, degli ormoni tiroidei (l’ipotiroidismo riduce la conversione dei carotenoidi in retinolo), il contenuto di zinco e di vitamina C ed E.
Quando gli apporti proteici sono deboli il β-carotene è convertito con difficoltà in vitamina A
Nel diabete mellito il β-carotene non è convertito in vitamina A.
La conversione in vitamina A di un carotenoide diminuisce quando l’apporto di carotenoidi aumenta e quando i livelli di vitamina A nell’organismo sono adeguati.
Nelle cellule dei vari tessuti il retinolo viene in parte ossidato a retinale, e parte di questo in acido retinico. Ciascuno di questi composti esplica funzioni peculiari.Il retinale e il retinolo sono interconvertibili, mentre l’acido retinico non può essere ridotto a retinale.
La carenza di vitamina A e di carotene si manifesta soprattutto nei tessuti che presentano un ricambio rapido, come quello epiteliale, che tende in tal caso ad ipercheratinizzarsi.
Se la vitamina A è carente, l’organismo si adatta, rallentandone il catabolismo.
L’esposizione a sostanze chimiche tossiche, alla diossina per esempio, può provocare un aumento di degradazione della vitamina A nel fegato, e di conseguenza un aumento del suo fabbisogno.
I sintomi di un’eventuale carenza sono: infezioni ricorrenti, infiammazione occhi (congiuntivite), incapacità di vedere bene al crepuscolo, cecità notturna, occhi stanchi dopo la lettura, otiti, secchezza mucosa della bocca e del sistema respiratorio, secchezza mucosa uro-genitale, bronchiti, pelle dura, acne, sintomi nervosi, problemi gengivali.
La carenza di vitamina A è preceduta, specialmente nei bambini, da raffreddori e disturbi respiratori.
Ad ogni respiro l’aria passa attraverso le fosse nasali rivestite di peli e muco che servono a trattenere particelle estranee. Tale rivestimento dipende dalla vitamina A ed in sua assenza l’inquinamento può invadere i polmoni.
Oltre a tutto questo si può avere perdita dell’udito e compromissione nelle capacità olfattive.
La vitamina A aiuta a sconfiggere le infezioni perché assicura la robustezza delle pareti cellulari ed aiuta ad impedire la penetrazione dei virus nelle cellule: in pratica è la nostra "armatura" della salute.
L’ aumento delle difese avviene sia in modo aspecifico, mantenendo in buono stato le superfici epiteliali e le mucose, sia stimolando ed intensificando molti processi immunitari, come l’induzione di attività antitumorali, il rafforzamento delle funzioni dei globuli bianchi e l’aumento della risposta anticorpale. La vitamina A, inoltre, aiuta il timo a mantenersi efficiente e ciò permette la produzione di cellule T.
È indispensabile per la sintesi proteica, essendo un fattore di crescita, soprattutto per ossa e denti. La vitamina A svolge un ruolo di primo piano nei processi biosintetici di molte glicoproteine, che regolano la differenziazione cellulare e l’espressione dei geni; per questo motivo è un fattore importante nello sviluppo e nella crescita dell’organismo umano.
La vitamina E accelera la circolazione del sangue e stabilizza la A, ma non solo: le vitamine A ed E necessitano una dell’altra, in quanto una impedisce che l’altra si ossidi.
Invece la vitamina A non deve essere mai assunta in contemporanea con la vitamina F in quanto hanno difficoltà di assimilazione, c’è una sorta di antagonismo. L’unico modo consiste nell’assumerle a 8 ore di distanza una dall’altra. La vitamina A è inoltre legata alla capacità di convertire il colesterolo in ormoni.
Quindi, la vitamina A è essenziale per la vista, ha un'azione protettiva nei confronti della pelle e delle mucose, delle ossa e denti denti, dei capelli e nell’ipercolesterolemia, favorisce i processi di crescita e contribuisce all'integrità del sistema immunitario.
Il fabbisogno giornaliero di vitamina A
In passato l’attività della vitamina A veniva misurata in Unità Internazionali (UI):
1 UI = 0,3 mcg di retinolo
oppure
1 UI = 0,6 mcg di β-carotene
Attualmente i livelli di assunzione di vitamina A vengono espressi in termini di retinolo equivalenti (RE) secondo le seguenti relazioni:
1 RE = 1 mcg di retinolo
oppure
1 RE = 6 mcg di caroteni provitaminici = 12 μg di altri carotenoidi non provitaminici
A causa del ridotto assorbimento intestinale e del metabolismo incompleto il rapporto carotenoide:vitamina sperimentato è pari a 6:1 (occorrono 6 microgrammi di carotene per ottenere un grammo di vitamina A) per i carotenoidi provitaminici, mentre è 12:1 per i carotenoidi non provitaminici.

Livello di assunzione giornaliera di vitamina A raccomandato dalla
S.I.N.U. (Società Italiana di Nutrizione Umana), revisione 1996.
Si consiglia un introito giornaliero di 700 RE per l’uomo e 600 RE per la donna. Nel caso di donne in gravidanza l'assunzione dovrebbe essere aumentata di circa 100 RE/die per un totale di 700 RE/die. In caso di allattamento la quantità di base va aumentata di circa 350 RE/die.
I bambini non dispongono ancora di depositi di vitamina A nel fegato per cui il loro approvvigionamento deve essere continuato.
Tra gli alimenti più ricchi di vitamina A ci sono il fegato e l'olio di fegato di merluzzo, le uova e i latticini; abbondanti quantità di carotenoidi sono presenti in frutta e ortaggi giallo-arancioni (pomodoro, carota, peperone, zucca, albicocca) e in alcuni ortaggi a foglia verde (lattuga, asparagi, broccoli).
L'assunzione di dosi di vitamina A ≥300 mg può determinare la comparsa di una sindrome tossica nota come ipervitaminosi A. I sintomi precoci comprendono secchezza e prurito cutaneo, desquamazione cutanea, dermatite eritematosa, alterazioni dell’accrescimento dei peli, perdita dei capelli, fessurazione delle labbra, cefalea, anoressia, edema, affaticamento, dolori muscolari anemia. Seguono vomito, capogiri, sonnolenza e sintomi neurologici vari.
Minore è l’età del soggetto, maggiori sono i rischi connessi.
Rispetto ai casi di tossicità acuta, quelli di tossicità cronica sono più frequenti in quanto è sufficiente assumere dosi giornaliere superiori alle capacità di immagazzinamento ed eliminazione del fegato sia tramite consumo frequente di alimenti ricchi in vitamina A sia per mezzo di integratori.
Per evitare ciò si consiglia di non superare l'assunzione di 9 mg/die per l’uomo e 7,5 mg/die per la donna. In gravidanza si consiglia di non superare i 6 mg/die in quanto più predisposte alla comparsa di sintomi da iperdosaggio. Dosi giornaliere superiori ai 30 mg/die sembra che possano avere un effetto teratogeno sul feto per cui si consiglia di non assumere mai integratori di vitamina A in gravidanza se non dietro prescrizione medica, la quale, tuttavia, non deve superare i 6 mg/die.
I caroteni non danno ipervitaminosi, l'effetto sfavorevole è stato osservato solo per dosi molto più elevate di quelle fisiologiche, ottenibili solo con preparati farmacologici od integratori (180mg/die); non solo non vi è alcuna evidenza invece che dosi fisiologiche di carotenoidi, quali quelle assunte con la dieta, siano dannose, ma la letteratura è assolutamente concorde sul fatto che questi, assieme a tutti gli elementi benefici presenti negli alimenti che li contengono, sono sicuramente in grado di proteggerci da malattie cardiovascolari e da diversi tipi di tumori.
La tossicità da carotenoidi è più contenuta in quanto il loro assorbimento è modesto e diminuisce ancora in caso di alte dosi.
Perché il carotene venga assorbito nell’intestino è richiesta la presenza di acidi biliari. Altri fattori possono inoltre favorire questo processo, come la presenza di grasso, proteine ed antiossidanti nel cibo ingerito, un adatto livello di enzimi pancreatici nell’intestino e l’integrità delle cellule della mucosa. L’efficienza dell’assorbimento del β-carotene oscilla tra il 40 e il 60%, e decresce all’aumentare della sua concentrazione.
Il carotene, se non è subito convertito dalle cellule della mucosa intestinale a vitamina A, viene incorporato nei chilomicroni (sfere formate da sostanze grasse), per essere trasportato nel sistema linfatico, e da qui riversato nel torrente sanguigno.
Di solito è trasportato nel plasma associato a lipoproteine a bassa densità (LDL); questo fa si che pazienti con elevati livelli plasmatici di colesterolo e LDL presentino alti valori di carotene nel siero.
I livelli sierici di caroteni dipendono dai quantitativi introdotti con l’alimentazione.
Il carotene viene immagazzinato nel fegato, nel tessuto adiposo, nelle ghiandole surrenali, nei testicoli, nelle ovaie e nella cute.
Il deposito nella cute determina una colorazione giallastra della stessa (carotenodermia), conseguenza di una eccessiva assunzione, alimentare o integrativa, di caroteni, e anche di una eventuale deficienza di fattori necessari alla loro trasformazione, quali l’ormone tiroideo, la vitamina C, lo zinco o le proteine.
Infatti i caroteni che non vengono trasformati in vitamina A, e che non reagiscono con i radicali liberi, raggiungono poi l’epidermide dove svolgono 2 ruoli dermatologici importanti: contribuiscono alla pigmentazione fisiologica della pelle anche in assenza di sole e la proteggono contro gli effetti nocivi dei raggi UV, aumentandone la soglia di foto tossicità.
Dunque, in caso di sovradosaggio si può verificare la comparsa di pigmentazione giallo-arancione della pelle e delle mucose, fenomeno che regredisce in poco tempo una volta che la dose di carotenoidi venga ridotta.
La protezione dagli effetti dannosi del sole
L’entità dei danni che il sole può causare varia in funzione di numerosi fattori sia individuali (tipi di pelle e di pigmento cutaneo, stato di salute età) che ambientali (orario e tempo di esposizione, stagione, altitudine, fenomeni di riflessione alla luce, ecc). Per questo ogni persona ha necessità di una protezione che si adatti alle sue particolari condizioni.
E’ necessario pertanto usare filtri (le cosiddette creme abbronzanti) che completino la protezione cutanea data dal filtro naturale costituito essenzialmente dai peli, dallo strato corneo, dall’acido urocanico e dalla melanina.
Recentemente sono stati condotti studi che hanno dimostrato la possibilità di difendersi dal sole per via sistemica, attraverso l’assunzione di sostanze in grado di cambiare la reazione avversa indotta dagli UV, attraverso una modificazione delle vie fotochimiche.
Le sostanze prese in esame sono alcuni farmaci, vitamine A, B6, C ed E e molti carotenoidi, in particolare il β-carotene. I carotenoidi rappresentano un gruppo di principi attivi che sembra svolgere un ruolo importante nel sistema antiossidante soprattutto della pelle.
Infatti, ad esempio, il β-carotene ha dimostrato di possedere una notevole attività protettiva nei confronti della foto-carcinogenesi indotta nel topo nudo nel quale le manifestazioni tumorali da UV si manifestavano in ritardo ed in numero inferiore rispetto ai controlli.
Nel 1993 è stata valutata l’azione protettiva del β-carotene nei confronti dei danni provocati dalle radiazioni UV, in uno studio in doppio ceco, contro placebo, su 20 donne, esposte alla luce in condizioni controllate.
Si osservò che nelle donne che assumevano il β-carotene diminuiva il rischio di scottatura aumentando il livello soglia per la comparsa di eritema.
Questi effetti sono stati confermati da ulteriori studi condotti con altri carotenoidi esplicanti e non-esplicanti attività vitaminica A, utilizzati sempre come dietetici.
D’altra parte i carotenoidi non svolgono sicuramente un’attività filtrante nei confronti degli UV e quindi la loro azione protettiva nei confronti della cute è dovuta sicuramente alla loro capacità di neutralizzare i radicali liberi.
E’ stato così dimostrato da diversi autori come l’esposizione protratta agli UV provochi la formazione di radicali liberi ed il conseguente aumento nel consumo di carotenoidi.
E’ stato inoltre dimostrato come la regolare assunzione di carotenoidi attraverso la dieta incrementi di circa il 50% l’attività foto-protettiva dei filtri solari applicati sulla cute aumentandone il loro fattore di protezione e riducendo la presenza nel sangue di radicali liberi.
Il loro assorbimento dalle fonti naturalmente disponibili è molto limitato: dalle carote si può assorbire in genere solo il 5% di β-carotene, a causa di una matrice complessa di polisaccaridi digeribili ed indigeribili e proteine; il licopene presente nei pomodori è più assimilabile se assunto sotto forma di succo o conserva rispetto al frutto fresco ed non trattato.
Un modo molto efficace per assimilare carotenoidi è quello di assumerli sotto forma di integratori alimentari.
Un efficace fotoprotettore dovrebbe agire contro gli effetti nocivi dei raggi UV, quali scottatura, immunosoppressione, discromie, fotoaging e tumori cutanei. Gran parte di questi effetti sono dovuti alla formazione di specie reattive dell’ ossigeno (ROS) ed all’assorbimento di fotoni; altri consistono nei danni nei confronti del sistema immunitario.
L’esperienza ha ormai dimostrato che gli integratori contenenti carotenoidi sono efficaci nel ridurre i danni causati dell’ossigeno singoletto, uno dei ROS più attivi, attraverso gli effetti dell’eritema e della scottatura solare.
Esistono, inoltre, numerosi studi sull’effetto protettivo dei carotenoidi. Recentemente sono stati studiati il licopene e l’α-carotene, che sono risultati ancora più attivi verso l’ossigeno singoletto rispetto al β-carotene. Tuttavia, le ultime tendenze in fatto di integratori “pre-sun” sono quelle di utilizzare un mix naturale di carotenoidi contenenti anche componenti minori ed in grado di agire in sinergia fra loro, piuttosto che basarsi solo sui singoli carotenoidi. Inoltre, questo consente anche di aumentare i dosaggi mantenendo una bassa attività provitaminica-A.
Ecco pertanto che, oltre al β-carotene, possiamo trovare integratori con carotenoidi come l’α-carotene ed il γ-carotene, estratti dal frutto della palma da olio (Elaeis Guinensis), il licopene, estratto dai pomodori, la luteina e la zeaxantina, estratte dai fiori di calendula, o la Polypodium leucotomos, un estratto di felce della America Centrale.
Ogni carotenoide ha le sue peculiarità: luteina e zeaxatina, ad esempio, non hanno funzione protettiva solo a livello cutaneo, ma proteggono anche l’occhio dagli effetti degenerativi della luce blu e degli UV sulla macula, consentendo una prevenzione nei confronti della degenerazione maculare legata all’età (DMLE).
L’effetto antiossidante dei carotenoidi può ridurre anche i danni indotti dall’esposizione agli UV sulle cellule di Langerhans. Tuttavia, alcuni fotoprotettori sistemici integrano la difesa delle cellule di Langerhans attraverso l’impiego di probiotici come il lattobacillo La 1 o, più recentemente, il bifidobatterio BB12. I probiotici sono definiti i microrganismi che, somministrati ed assorbiti in dosi adeguate, hanno effetto salutare per l’ospite (FAO/WHO 2001; 2002). Lo stato infiammatorio provocato dall’esposizione ai raggi UV si traduce in un aumento dell’interleuchina 10 ed in una diminuzione dell’attività delle cellule di Langerhans: la somministrazione di probiotici, prima e durante l’esposizione, rinnova l’attività di queste cellule nel presentare l’antigene.
La sinergia dei carotenoidi con i probiotici inizia dall’assorbimento: infatti, i carotenoidi sono liposolubili e possono essere assorbiti per via enteroepatica, laddove la caratteristica dei probiotici è quella di venire assorbiti a stomaco pieno, essendo resistenti ai succhi gastrici ed alla bile e potendo giungere nell’ intestino vivi e vitali, con elevata capacità di adesione alle pareti intestinali. Ciò permettere di assumere questi integratori a stomaco pieno, permettendone la massima efficacia.
L’attività fotoprotettiva del β-carotene si esprime sia in modo diretto che indiretto, per interazione con altri composti antiossidanti quali l’acido ascorbico e l’α-tocoferolo (vitamine C ed E) e altri carotenoidi.
La vitamina E, in particolare, è considerata il maggior antiossidante liposolubile a livello delle membrane cellulari.
Il β-carotene, assieme ad altri antiossidanti, inibisce le reazioni di perossidazione dei lipidi nella pelle e neutralizza i radicali liberi o le specie di ossigeno reattive (ROS) che si generano nell’organismo per esposizione ai raggi UV.
Per svolgere questa importante azione antiradicalica, essi devono essere però presenti nel luogo di formazione dei radicali in concentrazione adeguata.
La radiazione UV incrementa il numero di radicali liberi e forme reattive dell’ossigeno, come l’anione superossido, l’idroperossido e l’idrossile, che essendo molto reattive ed instabili (la loro vita dura alcune frazioni di secondo), intervengono in numerose reazioni biochimiche e processi metabolici, portando all’alterazione e, col tempo, all’invecchiamento precoce dell’epitelio non protetto.
I radicali liberi sono infatti implicati sia nel danno acuto da UV (scottatura solare), che nella fotocarcinogenesi e nel fotoinvecchiamento.
Per la loro azione fotoprotettiva e l’ottima compatibilità, ne viene raccomandato l’uso nelle fotodermatosi (per esempio causate da antibiotici o sulfamidici), durante l’assunzione di farmaci fototossici e nella prevenzione delle lesioni UV-indotte sulla pelle.
Un altro studio effettuato utilizzando un mix di carotenoidi, estratti dall’alga Dunaliella salina, con un dosaggio di 50 mg/die, ha dimostrato chiaramente che la pigmentazione della pelle, determinata dall’assunzione di carotenoidi, esercita una ulteriore protezione, poiché cambia la capacità riflettente della cute: meno luce viene assorbita nel tessuto ed una maggior quantità è riflessa dalla superficie cutanea.
Tale effetto è importante, specie se combinato con l’applicazione topica di filtri solari, la cui efficacia può essere addirittura, indirettamente, raddoppiata, come è stato evidenziato combinando la somministrazione orale di β-carotene con l’utilizzo topico di diossido di titanio.
Bisogna tenere ben presente che: i carotenoidi non sostituiscono i filtri solari, che devono essere sempre applicati nell’esposizione al sole, ma ne coadiuvano l’azione, provvedendo a fornire una protezione addizionale, soprattutto a quelle persone che necessitano di prodotti topici con fattori di protezione molto elevati.
Quel che è stato recentemente dimostrato è anche la loro capacità di modulare la melanogenesi sia in presenza di ipopigmentazioni cutanee quali la vitiligine che nelle iperpigmentazioni da farmaci o da estrogeni.
Infatti, nella vitiligine l’assunzione di carotenoidi assieme alla contemporanea esposizione a UVB di una determinata lunghezza d’onda sembrano in grado di stimolare i melanociti ad allungare i propri prolungamenti dendritici melanizzando i cheratinociti limitrofi alle aree ipopigmentate.
D’altra parte nelle iperpigmentazioni sempre i carotenoidi sembrano in grado di ridurre l’iperproduzione di melanina prodotta nelle zone a più alta intensità di colore.
Infine sempre se assunti un mese prima di recarsi al mare o in montagna questi complessi composti accelererebbero la produzione di melanina nelle persone normali oltre che nelle persone a rischio, facilitando la reazione melanogenetica che porta a formazione di melanina.
Bibliografia
1. B. TIRILLINI “Fondamenti di fitochimica” Ed. AKROS.
2. F. MEARELLI, A. GIOGLI “I CAROTENOIDI e la protezione dell’organismo” www.plantamedica.it
3. MAFFEI “ Biochimica Vegetale” Ed. PICCIN.
4. A. BRUNI “ Biologia Vegetale” Ed. JAPADRE.
5. G. MUSTICH “ Approfondimenti di chimica” Natural 1 Luglio – Agosto 2005 ANNO V – N°44.
6. R. LONGO “Alimentazione Ragionata” Natural 1 Gennaio – Febbraio 2006 ANNO VI – N° 49.
7. P. MORGANTI “I Carotenoidi quale fonte cutanea di benessere” www.mavicosmetics.it
8. M. MULTINEDDU “luteina e zeaxantina: gli antiossidanti per il benessere degli occhi” Dicembre 2006 www.lerboristeria.com